膜輔蛋白(CD46)
膜輔蛋白(membranecofactor protein,MCP)由Cole等(1985)應(yīng)用C3b親和層析在外周血**細(xì)胞上發(fā)現(xiàn)的一種膜蛋白�����。由于其奇特的電泳特征,起初被命名為gp45-70�,但進(jìn)一步研究發(fā)現(xiàn),它對(duì)I因子介導(dǎo)的對(duì)C3b和C4b的裂解有輔助活性�����,故更命為MCP�。鑒于MCP的多克隆抗體與促衷變因子(DAF)、H因子或CR1均不起反應(yīng)�����,從而認(rèn)為它是補(bǔ)體系統(tǒng)的一種新的調(diào)節(jié)蛋白�,在第四屆國際白細(xì)胞分型討論會(huì)上將其命名為CD46。
MCP為一單鏈穿膜糖蛋白����,分子量45-70kDa�����,屬于RCA基因簇的成員����。也通過GPI錨固定于細(xì)胞上�。MCP的細(xì)胞分布甚廣�����,包括粒細(xì)胞�、血小板、T細(xì)胞(Th��、Ts��、Tc)�、B細(xì)胞、NK細(xì)胞�、造血細(xì)胞系、成纖維細(xì)胞����、表皮細(xì)胞、內(nèi)皮細(xì)胞及星狀膠質(zhì)細(xì)胞等���。但不同類型的細(xì)胞上表達(dá)的數(shù)量有所不同����。外周血單個(gè)核細(xì)胞和粒細(xì)胞為1萬個(gè)/細(xì)胞,造血細(xì)胞系為2-5萬個(gè)/細(xì)胞�,Hela和Hep-2細(xì)胞分別為10萬個(gè)/細(xì)胞和25萬個(gè)/細(xì)胞。這種表達(dá)數(shù)量的差異���,可能反映了MCP在正常細(xì)胞和腫瘤細(xì)胞上不同的分化和活化狀態(tài)����。另外�,由于不同細(xì)胞上表達(dá)的MCP不同,因此可調(diào)節(jié)兩條激活途徑的C3轉(zhuǎn)化酶形成��。這一點(diǎn)尤其重要�,因?yàn)镃3慢速運(yùn)轉(zhuǎn)(tickover)的機(jī)制,能夠連續(xù)不斷的產(chǎn)生C3b與C4b��,有可能形成越來越多的C3轉(zhuǎn)化酶���。而由于大多數(shù)正常細(xì)胞上有高水平的MCP�,因此可保護(hù)正常細(xì)胞免遭補(bǔ)體介導(dǎo)的損傷�。相反,許多異物顆粒和致病微生物則缺乏MCP���,這樣沉積在它們表面的C3b便可得以保持其活化;且C3b與B因子結(jié)合的親和力高于與H因子結(jié)合的親和力,從而促進(jìn)c 3bBb復(fù)合物的形成����,導(dǎo)致在這些異物顆粒上補(bǔ)體有效地活化,*終將其破環(huán)**��。此外���,細(xì)胞表面唾液酸的含量與其結(jié)合H因子和B因子的親和力有關(guān)�,唾液酸含量較高的細(xì)胞與H因子的結(jié)合親和力超過與B因子的結(jié)合親和力��,唾液酸含量較低的細(xì)胞則與此相反���。許多**表面涶液酸的含量低于哺乳類動(dòng)物和人的細(xì)胞��,因此侵入機(jī)體后易同B因子結(jié)合而導(dǎo)致補(bǔ)體替代途徑的激活�����。
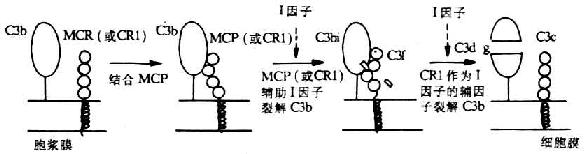
圖5-17 MCP輔助I因子裂解C3b的機(jī)理
MCP作為一種補(bǔ)體調(diào)節(jié)蛋白具有重要的生物學(xué)功能��。在低離子強(qiáng)度下�����,它可與C3b或C3bi結(jié)合�,但較CR1的親和力低。MCP的主要功能是其輔因子活性�����。即可與C3b或C4b結(jié)合而促進(jìn)I因子對(duì)C3b和C4b的裂解滅活(圖5-17)����,從而保護(hù)自身宿主細(xì)胞免遭補(bǔ)體介導(dǎo)的溶解破環(huán)。有人將MCP的這種作用稱為內(nèi)源性輔因子活性����。Seya等還發(fā)現(xiàn)MCP具有增強(qiáng)C3轉(zhuǎn)化酶的活性,尤其對(duì)替代途徑的C3轉(zhuǎn)化酶�����,但其生理意義尚不明了�����。CR1和H因子也是具有輔因子活性的補(bǔ)體調(diào)節(jié)蛋白�����,但H因子的這一活性僅為MCP的1/50。
人MCP的基因定位于第1號(hào)染色體長臂32區(qū)���,基因長度至少為43kb,含有14個(gè)外顯子和13個(gè)內(nèi)含子���。Seya等用從HSB-2T細(xì)胞純化獲得的MCP氨基末端設(shè)計(jì)了一個(gè)17mer的反義寡核苷酸探針����,以此從U937的cDNA文庫中篩選到一條長1.5kb的cDNA���。經(jīng)測序表明�����,該cDNA中含有一個(gè)43bp的5`端非編碼區(qū)和一個(gè)編碼384個(gè)氨基酸的開讀框���。其中前34個(gè)氨基酸為信號(hào)肽,后350個(gè)氨基酸為MCP的多肽鏈�,分子量為39kDa。在多肽鏈的前250個(gè)氨基酸中����,含有4個(gè)相鄰的CSR����。SCR后為一段富含絲氨酸�����、蘇氨酸和脯氨酸殘基的29個(gè)氨基酸片段(可能是高度O-連接的 糖基化位點(diǎn))����,再后依次為13個(gè)氨基酸的功能不明區(qū)、24個(gè)氨基酸的跨膜區(qū)����、105個(gè)氨基酸的胞漿錨和23個(gè)氨基酸構(gòu)成的胞漿尾部。大多數(shù)MCCP的變異局限在富含絲氨酸/蘇氨酸(S/T)區(qū)和胞漿尾部���。在核苷酸序列上MCP與DAF具有同源性����。
五���、H因子
H因子由Nilson等(1965)發(fā)現(xiàn)���,根據(jù)電泳位將其命名為β1H��,而Whaley和Ruddy則將其命名為C3b滅活劑加速因子?���,F(xiàn)已確定其為由1213個(gè)氨基酸組成的單鏈糖蛋白�����,分子量155kDa��,既有長桿狀部分�����,也有球形區(qū)域����。新近用透射電鏡檢查發(fā)現(xiàn)�����,H因子的影象為一長而柔順的分子��。伸長型分子長49.5nn�,橫截直徑為3.4mm但大多數(shù)不呈線型而呈折疊狀����,因此分子的實(shí)際長度僅為伸展型的一半����,但其構(gòu)象則呈多樣化。通過園二色譜分析表明��,H因子既無α螺旋也無β折疊����,借二硫鍵維持其功能活性的構(gòu)象。H因子的功能包括以下幾個(gè)方面:(1)為Ⅰ因子的輔助因子�����,可增加C4b對(duì)Ⅰ因子的敏感性�����。在無H因子時(shí)�,Ⅰ因子與C3b的結(jié)合呈絲狀;而在有H因子存在時(shí)��,I因子與C3b的結(jié)合變?yōu)閺澖z狀,同時(shí)與C3b的結(jié)合親和力增強(qiáng)(較無H因子時(shí)至少高15倍)����。H因子強(qiáng)化I因子的機(jī)理,可能是H因子與C3b結(jié)合后���,使C3b出現(xiàn)某些構(gòu)象變化�����,增加了與I因子結(jié)合的親和力��。H因子與C3b結(jié)合的活性部位存在于其N端35kDa部分。(2)加速C3轉(zhuǎn)化酶的衷變:H因子能將已同C3b結(jié)合的B因子或Bb從C3酶中逐出����,而使之失去酶活性。(3)阻止替代途徑中初始和放大C3轉(zhuǎn)化酶的形成�。已證實(shí)H因子和B因子在C3b上有同一結(jié)合部位,故H因子可同B因子或Bb競爭與C3b的結(jié)合��。在有H因子存在時(shí)�,B因子不易與C3(H2C)及C3b結(jié)合,因此不易形成C3(H2C)Bb或C 3bBb�。但H因子對(duì)固相上和液相中的C3b作用在差別。對(duì)液相中或結(jié)合于非激活劑固相上裂解��。而對(duì)于固定到激活劑(如酵母多糖等)表面的C3b,H因子則對(duì)C3b的親和力與B因子相當(dāng)�,二者競爭的結(jié)果,可形成部分C3轉(zhuǎn)化酶��,以保證替代途徑的活化���。有研究報(bào)道���,在細(xì)胞膜上能增強(qiáng)C3b對(duì)H因子親和力的化學(xué)成分是涎酸和肝素氨基多糖。由于大多數(shù)**表面缺乏涎酸��,因而這些**侵入機(jī)體后�����,可活化替代途徑��,有助于在早期對(duì)感染的控制�����。(4)對(duì)已與P因子或腎炎因子(NeF)結(jié)合形成穩(wěn)定的c 3bBbP或C 3bBbNeF�,H因子對(duì)它們也有一定的作用,但其效力要比CR1差得多。H因子對(duì)C5轉(zhuǎn)化酶(c 3bnBb或C 3bBb3b)的活性也有抑制作用����,并能與C5競爭結(jié)合C3b使C5不能裂解。此外����,近年發(fā)現(xiàn)H因子還可誘導(dǎo)單核細(xì)胞分泌IL-1參與**應(yīng)答的調(diào)節(jié)。
編碼H因子的基因定位于人的第1號(hào)染色體的長臂32區(qū)��,具有多態(tài)性��。已發(fā)現(xiàn)其有5個(gè)變異型�����,即FH1-5��。H因子的核苷酸序列已進(jìn)行了鑒定�,并推導(dǎo)出其全部氨基酸的**結(jié)構(gòu)�,含有20個(gè)SCR借此與C3b結(jié)合。H因子與MCP����、CR1、CR2、DAF及C4bp具有同源性���,共同屬于補(bǔ)體激活調(diào)節(jié)劑(RCA)基因家族的成員�����。
六����、I因子
I因子(舊稱C3bINA)為異源二聚體血清蛋白�,呈雙球狀結(jié)構(gòu),分子全長13nm���。其中小球(L鏈)4.9nm��,具有絲氨酸蛋白酶活性��;大球(H鏈)5.4nm可與C3b結(jié)合���。I因子的分子量為88kDa,重鏈50kDa�,輕鏈38kDa,鏈間以二硫鍵相連接����。I因子的生物學(xué)活性是��,在C4bp��、MCP���、H因子和CR1等輔助因子的協(xié)同下,將C4b裂解為C4d和C4c;使C3b裂解出C3f形成C3bi,后者再進(jìn)一步裂解為C3dg和C3c����,由此而控制補(bǔ)體系統(tǒng)的活化。
編碼入I因子的基因定位于第4號(hào)染色體上���。經(jīng)對(duì)I因子的cDNA序列分析發(fā)現(xiàn)��,其輕鏈為絲氨蛋白酶的活性區(qū)����,與糜蛋白酶�����、胰蛋白酶及彈性蛋白酶等有同源性���。而其重鏈?zhǔn)蔷哂懈缓腚装彼岬墓δ軈^(qū)����,與C8�、C9和低密度脂蛋白受體等具有同源性。I因子結(jié)構(gòu)基因的突變�����,可導(dǎo)致先天性I因子缺陷�����,此類患C3的過度消耗面引起反復(fù)感染和血管性水腫����。
七、過敏**滅活劑
過敏**滅活劑(AI)又稱血清羧肽酶N��,分子量310kDa���,由8條相同的多肽鏈組成���,每條分子量各36kDa�����。AI具有羧基肽酶的活性�����,可去除C4a�、C3a��、和C5a C末端的精氨酸殘基��,使這些片段喪失其過敏**活性����。
八、S蛋白
S蛋白(CP或S)為血清中一種α單鏈糖蛋白���,分子量83kDa�����。SP的主要調(diào)節(jié)作用是可與C5b~7的亞穩(wěn)態(tài)結(jié)合部位競爭靶細(xì)胞膜脂質(zhì)�����,通過形成親水性的SPC5b~7(簡寫為S5b~7)復(fù)合物�,而使C5b~7失去膜結(jié)合活性�。這樣,便可保護(hù)補(bǔ)體活化部位鄰近的細(xì)胞免遭偶然的攻擊��。這種親水性的SC5b~7還可集資與1個(gè)分子的C8和3個(gè)分子的C9結(jié)合����,分別形成SC5b~8和C5b~(9)3復(fù)合物,并C9聚合形成孔道��,從而可保護(hù)補(bǔ)體活化部位鄰近的細(xì)胞免于遭受補(bǔ)體的攻擊而損傷�����。SP與C8和C9的結(jié)合部位為這兩種分子中富含半胱氨酸的功能功能區(qū)����。電鏡下觀察,SC5b~(9)3復(fù)合物呈一楔形結(jié)構(gòu)��,SP位于楔形的寬部可掩蓋補(bǔ)體蛋白的疏水區(qū)���,從而封閉MAC的膜結(jié)合部位�。此外,2~3個(gè)分子的SP與C5b~7與C5b~8復(fù)合物的結(jié)合���,還可使這些復(fù)合物易溶�����,出現(xiàn)親水向疏水轉(zhuǎn)換��。SP也參與凝血過程�,通過干擾抗凝血酶Ⅲ對(duì)凝血酶的來活而保護(hù)凝血酶��。
編碼人SP基因定位于第17號(hào)染色體的長臂上���,其cDNA已克隆成功�����。經(jīng)過序列分析表明�,其與具有細(xì)胞粘附作用的玻璃粘連蛋白(vitronectin)的序列完全相同���,已證明二者屬同一蛋白�����。